Comodo Dragon
Systematic and Distribution
The Komodo dragon, Varanus komodoensis, was described for the first time by Major Peter A. Ouwen in 1912 (Auffenberg & Auffenberg 2002; Dunn 1927). This giant lizard species was placed in the genus Varanus, family Varanidae, order Squamata, Class Reptiles (Mattison 1992). Varanus salvadorii from Southern New Guinea and V. varius from Southeastern and Eastern Australia are believed to be the sister groups of V. komodoensis (King et al. 2002; Molnar 2004). The closest congeneric species occupying the same region is the Monitor lizard, V. salvator salvator (Auffenberg 1981).
Auffenberg (1981) reported that this species was called the “ora” by the local people of Komodo, Rinca, and West Manggarai. There are several local names described by Auffenberg (1981) from across its distribution in the Lesser Sunda region (see Table 1). The name “Komodo” was taken from the name of the island where the first specimens were taken, and which means “rats” (Dunn 1928). Table 1. Local names for Komodo dragon Local name Region Ora (also hora, lawora) Komodo, Rinca, West Manggarai Buaya darat (= land crocodile) Komodo, Rinca, West Manggarai Rugu (= Ora) Central Manggarai Si (also ugu; = lizard) Central Manggarai Lio (also ugu, = large monitor) Central Manggarai Pendugu (Grandfather of Ora) Central Manggarai Mbou (= Ora) Central Manggarai Source: Auffenberg 1981 Even though the Komodo dragon is the largest lizard in the world, this species has the smallest range of any large carnivore (King & Green 1999; Mattison 1992; Pough et al. 2001). In the early studies of the Komodo dragon, this species was found in the heart of the Lesser Sunda region on the islands of Komodo, Rinca, Padar, Gili Motang, Gili Dasami (also known as Nusa Kode), and the Western coast of Flores Island (Dunn 1928; Fig. 2.1). Five of the islands are within the boundary of Komodo National Park (PHKA 2000; Fig 1). In studies conducted after 1991, this species could not be found on Padar Island (Ciofi & de Boer 2004; Jessop et al. 2004; Sastrawan & Ciofi 2002). Present Status
Based on population surveys conducted by the park authority, there were approximately 2405 Komodo dragons living within Komodo National Park in 1998 (PHKA 2000, unpublished report). Ciofi and de Boer (2004) estimated that the population density of dragons on Flores was more than 60% lower than that reported for Komodo National Park (Table 2). Jessop et al. (2006 in press) estimated that the dragon population on Gili Motang Island has the lowest density of dragons of all the inhabited islands within the park (see Table 2).
The disappearance of resident Komodo dragons on Padar Island probably stemmed from the decline of Timor deer (Cervus timorensis) populations due to illegal hunting (Ciofi 1999; Ciofi & de Boer 2004). Pet and Subijanto (2001) reported that there were at least 3 cases (37.5 %) of deer hunting in Komodo National Park that had been sent to court during 2000-2001. Ciofi et al. (2002), Ciofi and de Boer (2004), and Primack (2004) stated that fragmentation and habitat disturbance as a result of the high population growth of humans are the main factors affecting Komodo dragon populations on Flores. Table 2. Average density of Varanus komodoensis in Komodo National Park (KNP) and Flores Island. Location Population density Number of sites KNP 1 / 33.25 km 4 Komodo 13.7 ± 1.67 / km Rinca 19.6 ± 3.13 / km Nusa Kode 5.1 ± 0.61 / km Gili Motang 3.2 ± 0.23 / km Flores 1 / 170.0 km 7 Source: Modified from Ciofi & de Boer 2004; Jessop et al. 2006 unpublished data. Even though the Komodo dragon is not threatened by the leather trade, like the congeneric Water Monitor (V. salvator), and is considered ‘dangerous’ to humans (King et al. 2002; Shine et al. 1996; Ellis 1998), hunting and trade in this species has been occurring for a long time. Since the 1930’s Komodo dragons and their eggs have been hunted illegally for zoo collections and for traditional medicine (Primack et al. 1988). Hien (2003) reported that the local people of Riung, Northwest Flores, claimed that they once illegally caught 50 live specimens of the Komodo dragon for a foreigner. However, the widespread hunting and trading of other reptiles including varanids, for their skins and for food (e.g. Shine et al. 1996) should be considered a potential threat to the Komodo dragon. The Komodo dragon is protected by international conventions; it is listed in Appendix I of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) and is classified by the International Union for the Conservation of Nature Resources (IUCN) as “vulnerable” due to its demographic decline and limited distribution (Ciofi et al. 2002). The World Conservation Monitoring Center (WCMC) also listed the Komodo dragon as “Rare” due to it’s restricted distribution (Ellis 1998). This species is protected in Indonesia under Act no. 5, on Conservation of Biological Resources and Their Ecosystems,1990,; and Government regulation no. 7, on Protected Wild Flora and Fauna, 1998. Primack (2004) included the Komodo dragon in his discussion on “conservation priorities” as he considered that this species met all the criteria of distinctiveness, endangerment, and utility3. Biologi, Ecology, and Reproduction
Compared to other species in the family Varanidae, Varanus komodoenis has an extremely large body size. Adults can reach up to 304 cm in total body length and weigh up to 81.5 kg (Jessop et al. 2006 unpublished data). Sastrawan and Ciofi (2002) recorded the largest Komodo dragon in their field study as being about 300 cm in length and weighing about 69 kg. Auffenberg (1981) recorded his largest specimen caught as being 260 cm in length and 54 kg in weight. This is similar in length to the largest specimens of its sister species, the Papuan monitor V. salvadorii, which reached about 265 cm in length (Horn 2004). Another large varanid species, occuring on the islands of Java, Bali and throughout Lesser Sunda region, V. salvator may reach up to 218 cm in length and weigh up to 25 kg (Gaulke & Horn 2004; Horne & Gaulke 2004).Hatchlings of V. komodoensis average 30.4 cm in total length and 0.08 kg in weight, and are considerably longer at hatching than other large varanid species (Auffenberg 1981). Ciofi (2004) recorded that hatchling V. komodoensis average 42 cm in total length and 0.10 kg in weight. Hatchling V. salvadorii possibly reach up to 49 cm in total length and 0.55 kg in weight (King & Green 1999).
Komodo dragons can be found from sea level up to about 800 meters in altitude, mainly in tropical dry and moist deciduous monsoon forests (Ciofi 2004). Indeed, this species is generally distributed over entire islands within KNP and western coastal on Flores but is rarely found above 500 meters (Auffenberg 1981). All varanids are insectivores or carnivores. Unlike other varanids that rely on smaller prey species, Komodo dragons are able to feed on larger vertebrate species, such as the Timor Deer (Cervus timorensis), water buffalo (Bubalus bubalis), or small wild boars (Sus scrofa). Adult Komodo dragons mostly rely on a sit-and-wait hunting strategy to catch their prey (Auffenberg 1981; Green et al. 1990; King & Green 1999; Pough et al. 2001). Hatchlings and juveniles, however, feed on a diverse diet of insects, small lizards, snakes and birds, and use more active hunting strategies than adults (Auffenberg 1981; Ciofi 1999; Mattison 1992). Female Komodo dragons are known to breed when they reach a body weight of around 20 kgs (King & Green 1999). Females begin nesting in August, as determined by the presence of recent digging activity on the nest, or by repeated observation of individuals in association with the nest during the nesting period. The nesting period is from August through November, with egg deposition occurring in September (Ciofi 1999; Jessop et al. 2004). Up to 38 hatchlings will emerge from the nest at the beginning of the dry season (Auffenberg 1981; Jessop et al. 2006 unpublished data). This early life-stage is dominated by high mortality. Cannibalism among Komodo dragons has been observed and was calculated to comprise 8% of adult dragon scats (Auffenberg 1981). Cannibalism has also been recorded in other Varanids, such as V. griseus, V. gouldii and V. gigantheus (King & Green 1999). Biology and Ecology of the Juveniles
Generally, mortality in lizards is highest during the first phase of their life because they are more vulnerable to predation (Mattison 1992). Poor maternal body condition and stress can decrease the dispersal tendency of juveniles (Meylan et al. 2002). Typically, natural populations show substantial variation, in locomotor performance and body size, which is related to offspring survivorship (Clobert et al. 2000). Thus, the growth and survivorship of offspring in reptiles is greatly affected by the animal’s environment (i.e. Brockelman 1975; Gans & Pough 1982).
Incubation temperature is known to contribute greatly to the quality of emerging hatchlings and affects their survivorship. Cold temperature during incubation can negatively effect hatchlings and hot temperatures can positively effect hatchlings (Elphick & Shine 1998; Phillips & Packard 1994; Qualls & Andrews 1998). This pattern is complex, however, as higher incubation temperature will result in earlier hatching but produce lighter and smaller hatchlings than lower temperature, which produce larger offspring that tend to survive better (King & Green 1999). Du and Ji (2003) reported that moderate temperatures produced optimum size, locomotor ability and success of hatchlings in soft-shelled turtle, while Ji and Du (2000) reported a similar pattern in colubrid snakes. In a further study on lizards, the only discernible influence on juvenile phenotypes was their rearing environment (Qualls & Shine, 2000). Brockelman (1975) found that a wide variety of factors can affect optimal body size and the ability to process energy effectively, and these were also affected by the process of competition, which the offspring must face before and during maturity. Most juvenile reptiles leave the natal area in which they were born and move into new habitats that are not already occupied or to avoid cannibalism by adults (Pough et al. 2001). Greenwood (1984) noted that natal dispersal among juveniles is also considered as a mechanism to avoid future inbreeding. Heatwole (1976) and Sarno et al. (2003) described natal dispersal as being driven by the competition for food resources and territories with adult.Dispersal is a mechanism for survival and is a consequence of permanent movement away from the natal site (Brown & Downhower 1988). Animals will exploit available resources once they are out of their natal sites (Greenwood & Swingland 1984). Pelletier et al. (2003) described how immature turtles immediately swam towards the ocean and steadily traveled long distances once released into the water. Hatchling of varanids tend to climb trees and spend most of their time in the upper strata of trees once they emerged from the underground nests (Auffenberg 1981; Bohme et al. 2004; Ciofi 2004; King & Green 1999). Previous Ecological Studies
The first scientific study on the Komodo dragon was conducted by Major Peter A. Ouwen, director of the Zoological Museum in Buitenzorg (now Bogor), Java, in 1912 (Auffenberg & Auffenberg 2002). Ouwen described the species for the first time. Later, Dunn (1927 & 1928) conducted the first significant observations and gave the initial information on the habitat and distribution of Komodo dragons. Auffenberg (1981) contributed to the first comprehensive study on the ecology and behavior of this species. On the basis of 13 months of field observations, Auffenberg (1981) provided the base line information on aspects of the Komodo dragon’s ecology and behavior.
The most recent field study on the Komodo dragon was conducted by Jessop et al. (2006 unpublished data). Since 2002, they undertook a broad scale investigation in the ecology and demography of the Komodo dragon, which has provided important information and instigated a program to monitor population trends. Although numerous field studies on the ecology of Komodo dragons have been conducted, yet there are no detailed studies on the ecology on juvenile Komodo dragons (Auffenberg & Auffenberg 2002). Walsh et al. (2002) studied growth in juvenile Komodo dragons, whilst Lemm (2004) reported a relationship between growth and nutritional treatment in captivity. Field and captive studies on this creature, including work on growth, chromosomes, physiology, genetics, ecology and social behaviour, parasites, microbiology, conservation and management, have provided valuable information to science and to ensure effective management of the species (Ciofi et al. 2002). The long-term management and conservation objectives underpinning survival of this species will be to maintain a genetically viable, self-sustaining, and free-living Komodo dragon population. Information on reproduction and broad scale ecology of this species is needed to support the management authority responsible to protect it, and, as pointed out by Jessop et al. (2004), information on offspring survivorship are vital for management planning to ensure the maintenance of this unique species in the wild. |
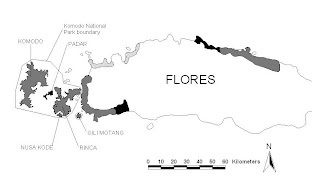
Distribution of Varanus komodoensis. Grey areas represent the curre distribution; black areas represent areas that were identified as part of the distribution by Auffenberg (1981); hatched areas indicate where V. komodoensis have been reported by villagers. Source: Redrawn from Sastrawan & Ciofi 2002.
Morales, Alex (2006-12-20). "Komodo Dragons, World's Largest Lizards, Have Virgin Births". Bloomberg Television. Retrieved 2008-03-28.
Henderson, Mark (2006-12-21). "Wise men testify to Dragon's virgin birth". The Times (London). Retrieved 2007-11-26. Komodo dragons hatch with no male involved. MSNBC. Retrieved 2008-02-12.^ a b "Virgin births for giant lizards". BBC News. 2006-12-20. Retrieved 2008-03-13. Strange but True: Komodo Dragons Show that "Virgin Births Are Possible: Scientific American". Scientific American. Retrieved 2008-03-24. Watts PC, Buley KR, Sanderson S, Boardman W, Ciofi C, Gibson R (Dec 2006). "Parthenogenesis in Komodo Dragons". Nature 444 (7122): 1021–2. doi:10.1038/4441021a. ISSN 0028-0836. PMID 17183308. |
